.jpg)
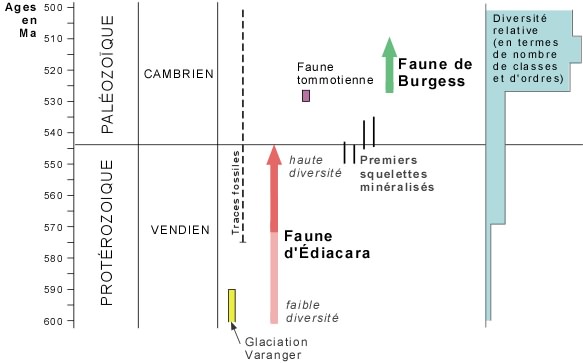
Une des périodes clé du développement de la vie sur terre se situe entre -600 et -520 Ma. C'est là que la diversité biologique a littéralement explosé.
On peut considérer que cette explosion de la diversité s'est faite en deux pulsations: un moment préparatoire, la faune d'Édiacara, puis la grande explosion, ce que certains appellent le "big bang" de la vie, représentée par la faune des schistes de Burgess.
Avant d'examiner chacune de ces trois faunes et voir ce qu'elles nous enseignent, il faut ouvrir une courte parenthèse sur la façon dont les paléontologues de la première moitié du 20e siècle appréhendaient ces nouvelles faunes.
Ces paléontologues avaient été profondément marqués par la pensée de Darwin sur l'évolution. Que la vie avait évolué progressivement d'une manière linéaire, selon la sélection naturelle, du plus simple au plus complexe, était à toutes fins pratiques devenu un fait acquis. On ne peut d'ailleurs leur en tenir rigueur, car ils travaillaient le plus souvent sur des groupes bien définis et sur des échelles de temps très courtes; ils pouvaient effectivement, à l'intérieur des groupes qu'ils étudiaient, établir des lignées évolutives où se reconnaissaient bien ancêtres et descendants. Cette façon de voir avait une implication très directe sur la méthode paléontologique: chaque organisme fossile avait un ancêtre qui devait être moins complexe - on disait moins évolué - et des descendants qui eux se devaient d'être plus "évolués". En pratique, lorsqu'on trouvait une forme énigmatique très ancienne dont on n'arrivait pas à bien saisir l'affinité biologique, on tentait de trouver à qui elle pouvait bien avoir donné naissance et on pouvait ensuite la classer dans ce groupe en disant voilà l'ancêtre.
Le tableau qui suit présente la répartition dans le temps et la diversité relative, selon les données les plus récentes, des faunes dont nous discuterons. Il vous aidera à mieux suivre la discussion. On a ici un intervalle de 70 millions d'années parmi les plus importants en ce qui touche l'évolution de la vie.
La faune d'Édiacara
La faune d'Édiacara est apparue il y a quelques 600 Ma et une grande partie de ses éléments est brusquement disparue 56 millions d'années plus tard, à - 544 Ma. Certains considèrent cette première faune comme correspondant à l'apparition des métazoaires [= organismes pluricellulaires, c'est-à-dire ceux qui possèdent des cellules diversement spécialisées, soit pour l'absorbtion des nutriments, le transport de diverses substances, la reproduction, etc.; par opposition à protozoaires, organismes dont les cellules remplissent toutes les mêmes fonctions, sans distinction].
Elle a d'abord été découverte dans les monts Édiacara en Australie, de là son nom. Par la suite, on a découvert une vingtaine de sites répartis sur les cinq continents. Ce point est important à signaler: il ne s'agit pas d'un assemblage d'organismes qui est apparu en un seul lieu particulier et qui y aurait été confiné, à cause de conditions spéciales à cet endroit, mais une faune répartie à la surface du globe, une faune qui représente une véritable étape dans le développement de la vie.
Un autre caractère très important de la faune d'Édiacara est qu'elle est composée en grande partie d'organismes à corps mous, c'est-à-dire des organismes sans squelette minéralisé. C'est donc dire que tout ce qu'on retrouve, c'est l'empreinte de l'animal sur le sédiment, et non l'animal fossilisé; ceci implique que l'interprétation joue un rôle important dans la compréhension de cette faune.
Ci-dessous quelques formes qu'on y trouve, représentées selon l'iconographie traditionelle. En fait, il s'agit de dessins interprétatifs qui datent du début des années 80. L'interprétation traditionnelle de cette faune est de considérer ses membres comme des représentants primitifs de groupes plus jeunes, essentiellement des membres de l'embranchement des cnidaires (coraux mous et médusoïdes), des annélides (vers segmentés) ou des arthropodes. Ces exemples montrent ici des organismes ressemblant, soit ŕ des méduses (a), soit ŕ d'autres cnidaires modernes comme les coraux mous (b), ou encore ŕ des arthropodes nus ou ŕ des vers annélides (c). Par contre, (d et e) ne ressemblent ŕ aucun animal connu.
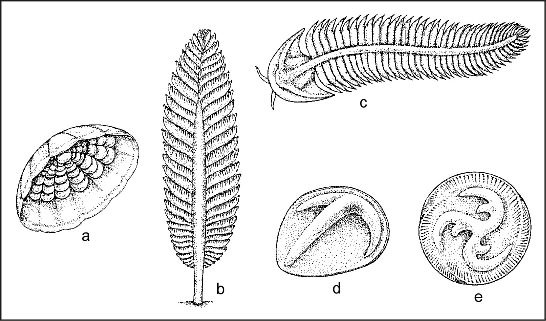
On a donné le nom de vendobiotes ŕ ce groupe d'organismes énigmatiques (de la période de temps où cette faune a vécu, le Vendien, intervalle de temps de la fin du Protérozoïque). [Dessins extraits de Cloud, P., 1983, La Biosphère. Pour la Science, novembre 1983, numéro spécial 73].
 | Les vendobiotes d'Édiacara en images |
Ci-dessous, une reconstitution de la communauté des vendobiotes sur le fond marin telle que conçue par l'interprétation traditionnelle. [Selon une vitrine du Smithsonian Museum ŕ Washington. Photo: P.-A. Bourque, 1995].

Un spécialiste de l'étude des traces fossiles, Adolph Seilacher des Universités de Tubingen (Allemagne) et de Yale (USA), a proposé une façon tout à fait différente d'interpréter cette faune. En utilisant les principes de la morphologie fonctionnelle, il a conclu que les vendobiotes n'ont pu fonctionner comme leurs supposés équivalents modernes, malgré une certaine similitude superficielle de leur apparence extérieure. Ils appartiendraient tous plutôt à un même groupe taxinomique, dans la mesure où ils ne représentent que des variations sur un même plan d'organisation anatomique: une forme aplatie divisée en parties formant un matelassage, constituant peut-être un squelette hydraulique, un peu à la manière d'un matelas pneumatique.

Ainsi, sur la rangée du haut, on a des formes à croissance unipolaire, c'est-à-dire dont la croissance part d'un seul point: ce sont les coraux mous et les vers annélides ou arthropodes nus de l'interprétation traditionelle. Sur la rangée du milieu, des formes à croissance bipolaire de part et d'autre d'une ligne centrale. Et sur la rangée du bas, des formes à croissance radiaire, à partir d'un point central; ce sont les méduses et les disques de l'interprétation précédente.
Dans la mesure où cette organisation ne ressemble à aucune de celles que nous connaissons actuellement, Seilacher a proposé que les vendobiotes constituent une expérience totalement distincte dans l'histoire de la vie. Il voit ces organismes comme des amas de cellules formant une sorte de matelas pneumatique protoplasmique étalé sur les fonds marins et absorbant passivement les nutriments du milieu, possiblement avec l'aide de symbiotes algaires. Cette expérience aurait finalement échouer, constituant une extinction précambrienne.
Le paléontologue Bruce Runnegar de l'Université de Californie continue pour sa part de considérer les vendobiotes comme les ancêtres de cnidaires comme les coraux mous et les médusoïdes, ou de certaines grandes algues marines, ou encore de vers ou d'arthropodes.
Il s'est fait beaucoup de travaux de recherches sur la faune d'Édiacara ces dernières années. Nous avons appris, entre autres, que la faune d'Édiacara n'est pas monolithique.
Il reste encore pas mal de chemin à faire avant que l'on comprenne bien cette faune d'Édiacara. Mais on peut dire d'ores et déjà qu'à côté de cette faune classique que sont les vendobionts qui n'étaient peut-être pas des métazoaires, il y avait des organismes actifs, certainement métazoaires, qui remobilisaient les sédiments et une faune coquillère à squelette minéralisé.
http://www.ucmp.berkeley.edu/vendian/critters.html
Un petit bout de texte et d'autres illustrations de la faune d'Édiacara avec descriptions de quelques formes.
La faune tommotienne
Une étape importante de l'évolution aprčs l'apparition des métazoaires est la minéralisation des squelettes. De ce point de vue, la faune tommotienne a longtemps été considérée comme faune clé. Elle tire son nom de Tommot, une petite ville de Sibérie. Comme la faune d'Édiacara, elle est aussi reconnue un peu partout au monde. Elle est apparue il y a 530 Ma et fut de courte durée (quelques millions d'années seulement). Jusqu'ŕ ces derničres années, on la considérait comme la premičre faune dotée de parties dures. Elle contient certains organismes d'aspect moderne, mais la plupart consiste en de minuscules lames, tubes, aiguilles, calottes ou coupoles d'affinités incertaines comme illustré ci-dessous (A ŕ C).
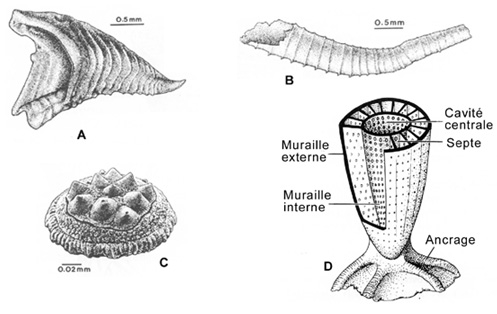
Les paléontologues l'appellent la "faune ŕ petites coquilles". Il s'agit peut-ętre simplement de fragments épars des premiers types de coquilles encore imparfaitement constituées. Peut-ętre que ces pičces recouvraient des animaux bien connus et que ceux-ci développčrent plus tard des coquilles plus élaborées constituant leur signature classique dans les archives fossiles. Mais peut-ętre que la plupart des étrangetés tommotiennes représentent des organismes uniques en leur genre qui sont apparus précocement et qui sont disparus rapidement.
Cependant, la découverte récente d'organismes à squelette minéralisé, dans des couches de la fin du Précambrien et du début du Cambrien (entre -550 et -540 Ma, donc en partie contemporaines de la faune d'Édiacara), reporte l'apparition de la minéralisation des squelettes antérieurement à la faune tommotienne.
La faune tommotienne est importante ŕ un autre titre: elle marque aussi l'apparition d'un groupe d'organismes appartenant fort probablement ŕ l'embranchement des éponges (Porifera) et qui n'a vécu que jusqu'ŕ la fin du Cambrien, les archaeocyathes (D). Ces derniers représentent les premiers bioconstructeurs qui ont édifié, en association avec des communautés microbiennes calcifiantes (calcimicrobes), des masses organiques de 1 ŕ 2 mčtres de hauteur par quelques mčtres de diamčtre sur les fonds marins. Ces masses sont les précurseurs de cet écosystčme récifal complexe qui s'est développé dčs la fin de l'Ordovicien (voir rubrique suivante).
Jusqu'ŕ tout récemment, les évolutionnistes considéraient qu'il y avait un "trou" important entre faune d'Édiacara et faune tommotienne (voir par exemple Gould, 1991, La vie est belle, Seuil) indiquant une décimation importante ŕ la fin du Précambrien. On réalise aujourd'hui que, si les vendobiotes semblent effectivement disparus, plusieurs autres organismes, dont les responsables des traces fossiles de remaniement et la faune coquillčre, constituent un pont entre faunes d'Édiacara et tommotienne.
La faune de Burgess
La troisième faune de cette période clé de l'évolution de la vie est la faune du Schiste de Burgess, la faune du "big bang". Par rapport aux deux faunes précédentes, elle constitue effectivement une véritable explosion, en termes de diversité et de complexité des organismes qui la constituent.
Cette faune, d'abord découverte dans le Schiste de Burgess dans le Parc national de Yoho, en Colombie-Britannique, a été retrouvée dans plusieurs parties du monde: Groenland, Europe, Chine, Australie et ailleurs en Amérique du Nord. Il s'agit donc d'une faune de répartition mondiale. Elle est apparue il y a 528 Ma et disparue brusquement à -510 Ma, représentant un intervalle de temps de presque 20 millions d'années.
Le Schiste de Burgess, comme tout schiste, résulte de l'induration d'une boue. Ce qui distingue cependant ce schiste des autres schistes, une roche très abondante à la surface du globe, c'est qu'il contient une faune fossile unique dans toute l'histoire de la vie sur Terre. Les animaux du Burgess vivaient probablement sur des talus de boue accumulés au pied d'une muraille massive presque verticale. Ce genre d'habitat ne constitue pas une situation exceptionnelle, mais tout à fait ordinaire; on ne peut donc invoquer de particularité écologique pour expliquer la singularité de la faune de Burgess. Ce qui est exceptionnel, c'est la conservation des parties molles de plusieurs de ces organismes qui s'est sans doute faite à la faveur d'un enfouissement rapide causé par des coulées de boue. L'absence d'oxygène et de nécrophages dans le milieu d'enfouissement expliqueraient la conservation.
Le site fossilifère du Schiste de Burgess fut découvert en 1909 par le paléontologue américain Charles Doolittle Walcott, alors secrétaire de la Smithsonian Institution de Washington. Il consacra une bonne partie de sa carrière de chercheur à la description de cette faune, jusqu'à sa mort en 1927. Il fallut attendre la fin des années 1960 pour que Harry Whittington de l'Université de Cambridge (Angleterre), en collaboration avec Simon Conway Morris et Derek Briggs, procède, sous l'impulsion de la Commission géologique du Canada, à une réévaluation en profondeur de cette faune.
Walcott était un spécialiste des faunes cambriennes et des trilobites en particulier. En accord avec les vues de son époque qui voulaient qu'une faune cambrienne soit l'embryon (donc une faune primitive) des faunes plus jeunes, il lui fallait classer chaque organisme dans un groupe connu. La force de l'équipe de Whittington a été de sortir de cette ligne de pensée et de reconnaître que plusieurs formes représentent des plans d'organisation anatomique inconnue aujourd'hui.
La somme actuelle des organismes décrits par l'équipe de Whittington se chiffre à 120 genres. Parmi ceux-ci, 33 genres sont des organismes ordinaires, appartenant à des embranchements que l'on connaît bien, et qui possèdent un exosquelette (éponges, algues, brachiopodes, trilobites, échinodermes, mollusques). Cependant, on dénombre 87 genres d'organismes à corps mou (c'est-à-dire sans squelette), ce qui fait l'unicité de cette faune. La présence d'organismes qui représentent des formes qui ne se retrouveront plus par la suite dans les temps géologiques est aussi unique. Entre autres, huit genres représentent autant d'embranchements qui n'ont pas survécu. Chez les arthropodes, on trouve 24 types fondamentaux d'organisation, alors que seulement 4 ont persisté. Actuellement, on a classé comme "animaux divers" ou "problematica", près de 40 formes, parmi lesquelles la probabilité de découvrir de nouveaux types d'organisation sont fortes.
Voici quelques exemples des trouvailles du Schiste de Burgess qui représentent chacune des essais infructueux d'embranchements (phyla). Tous ces dessins sont de Marianne Collins dans Gould, S.J., 1991, La vie est belle, Editions du Seuil, sauf celui d'Hallucigenia par Ramsköld (1992).





Parmi ces bêtes étranges, il en est une qui a donné de sérieux maux de tête à Simon Conway-Morris, ce qui explique le nom que ce dernier lui a donné: Hallucigenia. Tous les spécimens du site de Burgess en Colombie-Britannique examinés par Conway-Morris lui ont permis de faire la reconstitution suivante: un animal à corps allongé avec, d'un côté 7 paires d'épines droites, rigides et non-articulées, sauf à la jonction du corps, et de l'autre côté une seule rangée de 7 appendices flexibles. Conway-Morris n'avait donc d'autre choix que de conclure que l'animal devait se déplacer sur ses paires d'épines, comme sur des échasses. Imaginez un animal qui se déplace sur des échasses rigides pointues dans de la boue, ce qui était le substrat où vivaient les animaux du Burgess. Hallucinant!

Ce n'est qu'au début des années 90 qu'on a découvert, en Chine, des spécimens beaucoup mieux conservés que ceux du Schiste de Burgess, montrant en fait que les appendices flexibles que Conway-Morris voyait sur le dos de l'animal se trouvaient par paires (Ramsköld, L., 1992. The second leg row of Hallucigenia discovered. Lethaia, v. 25, p. 221-224). Ces appendices étaient certainement mieux adaptées à la marche dans la boue que les échasses de Conway-Morris.

Cette découverte venait aussi résoudre une des énigmes de la faune de Burgess: ainsi orienté, Hallucigenia s'apparente à des organismes connus, les onychophores, animaux de la forêt pluviale tropicale et tempérée (Ramskold, L. et X.-G. Hou, 1991. New early Cambrian animal and onychophoran affinities of engimatic metozoans. Nature, v. 351, p. 225-228).
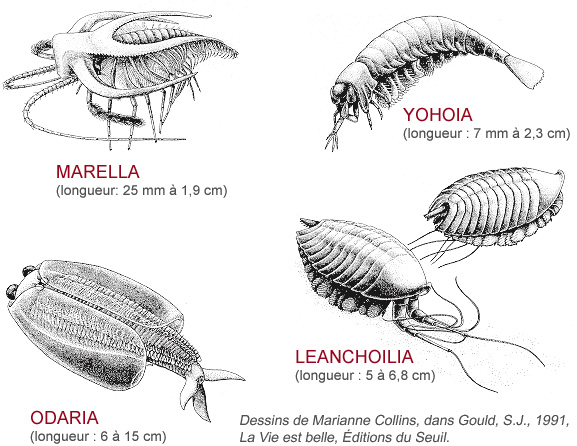
Outre ces étrangetés dont on ne connaît pas l'affinité, il y eut aussi des essais infructueux dans le groupe des arthropodes où seulement 4 essais sur 24 ont persisté. Voici quatre exemples qui portent toutes les caractéristiques fondamentales de l'embranchement des arthropodes. Elles font partie de ces 20 types d'organisation qui n'ont pas réussi.
Finalement, il faut signaler un tout petit animal, le tout premier chordé, trouvé dans le Schiste de Burgess. Notre ancêtre. S'il n'avait survécu à la décimation du stock initial de Burgess, nous ne serions peut-être pas là!

La grande conclusion à laquelle sont arrivés Whittington et ses collaborateurs est que la faune du Schiste de Burgess montre une extraordinaire différence entre la vie actuelle et celle d'un lointain passé: avec un beaucoup plus petit nombre d'espèces, le Schiste de Burgess présente une diversité des plans d'organisation anatomique bien plus grande que la gamme que l'on peut observer actuellement dans le monde entier. En un instant géologique, au milieu du Cambrien, presque tous les embranchements modernes ont fait leur apparition, en même temps qu'une vaste gamme de formes animales qui sont autant d'expériences anatomiques, mais qui ne survivront pas très longtemps. Les 500 Ma suivants n'ont vu naître aucun embranchement (sauf peut-être les bryozoaires), seulement des variantes sur des modèles de base établis.
L'étude de cette faune nous enseigne que l'histoire de la vie multicellulaire a été dominée par la décimation d'un vaste stock initial, qui s'était constitué en peu de temps lors de l'explosion cambrienne. L'histoire des 500 derniers Ma a été caractérisée par la restriction de la disparité, suivie de la prolifération de quelques types d'organisation stéréotypée, et non pas par l'expansion générale de la gamme des plans anatomiques et d'un accroissement de la complexité, comme le voudrait notre conception de l'évolution selon un cône de diversité croissante.
GOULD, S.J.,1991, La vie est belle, Editions du Seuil. L'aventure merveilleuse de la redécouverte de la faune de Burgess par l'équipe de Whittington, étape par étape, et les leçons qu'on en tire sur notre conception de l'évolution. Passionnant.
NASH, J. Madeleine, 1995, When life exploded. Un superbe article du TIME Magazine, décembre, 1995, qui présente un excellent résumé bien documenté des récentes découvertes, des implications et des questions qu'elles suscitent en ce qui touche l'évolution dans cette période cruciale de la fin du Précambrien - début du Cambrien. Du grand journalisme scientifique!
http://www.burgess-shale.bc.ca Le site officiel de la Fondation Burgess.
http://www.geo.ucalgary.ca/~macrae/Burgess_Shale/ Le point de départ pour tout ce que vous voulez savoir sur le Schiste de Burgess, avec des illustrations, des photos, plusieurs liens, etc.
 | Retour au plan 4.3.3 |  |